All published articles of this journal are available on ScienceDirect.

Marine-Derived Proteins: Emerging Nutritional Strategies for Optimizing Athletic Performance
Abstract
Marine-derived proteins and peptides are emerging as an exciting area of interest in terms of their potential for delivering improved performance through functional nutrition. Marine proteins, which have a good balance of essential amino acids, are easily digested and contain bioactive peptides that have antioxidant, anti-inflammatory, and metabolism-modulating properties. Recent scientific studies have found that marine protein can contribute to muscle building through the strengthening of connective tissues via collagen. Marine bioactive peptides are being studied to enhance vascular functions, reduce oxidative stress levels, and increase endurance and metabolism. This review outlines the existing literature on the biochemical properties, physiological pathways, and potential applications of marine proteins and peptides in sports nutrition, highlighting their increasing role in delivering enhanced performance through functional nutrition.
1. INTRODUCTION
Marine environments occupy more than 70% of the Earth’s surface and provide an unprecedented level of biological diversity from microscopic phytoplankton through larger marine mammals. The capacity of the organisms in these environments to sustain life in so-called harsh environments with high levels of salinity, pressure, and temperature gradients has played a role in the development of proteins/peptides with unique structures and superior functional properties insofar as their molecular diversity and levels of activity are concerned. In this respect, bioactive compounds from marine organisms offer higher levels and greater promise for applications in sports [1, 2]. Proteins are very significant in sports nutrition as a source of energy and essential amino acids, which are important in muscle protein synthesis. Bioactive peptides are compounds found in dietary proteins, consisting of short sequences of amino acids (typically 3-20) that lack biological activity in their native peptide form [1]. These peptides can exert their influence on exercise-related biological processes when released. These biological processes depend on amino acid sequences that produce differences in molecular weight. There has been a significant increase in research on marine peptides derived from fish, seafood, shellfish, seaweed, microalgae, or other by-products of marine processing. With regard to sports, the applicability of these marine peptides can be beneficial for reducing oxidative stress associated with high-intensity exercise, inhibiting exercise-induced muscle injury, improving exercise-induced inflammation, and supporting mechanisms of exercise recovery, which are crucial for maintaining consistent exercise performance. Due to their high bioavailability and safety profile, often observed with marine peptides, their use as sports foods or nutraceuticals is particularly ideal [1, 3]. However, there have been breakthroughs that have ensured the capability to identify and analyze the potential marine peptides. Enzymatic hydrolysis is more desirable compared to other hydrolysis processes due to its efficacy, selectivity, and absence of chemical residues. This makes it suitable for application in the food industry [4-7].
The trend in the use of performance boosters is being reflected in a combination of animal, plant, sea, and dairy-based proteins. Of these, the use of marine proteins is gaining traction due to their content of amino acids. A side-by-side view of these nutritional trends is portrayed in Fig. (1). With regard to sports science, the general meaning of “optimizing athletic performance” refers to improvements across a range of physiological and purposeful domains that ultimately influence performance outcomes. The exact domains that are commonly comprised in a definition tend to be: (i) muscular strength that relate to the athlete’s ability to exert force during resistance training; (ii) endurance potential that is frequently designated by VO2 max and time to exhaustion; (iii) recovery potential that is defined by reduced muscle damage and inflammation; (iv) and metabolic potential that is designated by glycogen and mitochondrial function and energy efficiency. This review targets the rapidly developing use of marine proteins and peptides as dietary approaches for sports performance and exercise recovery. In making connections between progress in marine biotechnology and foundational principles in exercise physiology/sports nutrition, this review seeks to offer a focused view into the relevance of marine bioactive compounds to exercise adaptation and recovery or performance outcomes generally that have not received the same level of attention within more general nutraceutical review publications or discussions.
Nutritional strategies for sports performance emphasize the importance of proteins obtained from marine sources.
1.1. Marine-Sourced Peptides and Proteins: Their Sources and Characteristics
Marine proteins, which carry several biochemical and physiological benefits, are known for being easily digested and quickly absorbed by the body. This means that a surge of amino acids can be delivered into the muscles almost immediately after a training session. Furthermore, the amalgamation of amino acids in marine proteins is considered to be optimally balanced, containing great levels of branched-chain amino acids that interact well with muscle protein synthesis signaling cascades like mTOR. In addition, marine proteins, which encompass bioactive peptides, exhibit anti-inflammatory, vasodilatory, and antioxidant effects that may reduce oxidative stress that arises during high-intensity physical activities. In addition to these benefits, marine proteins also appear to contain omega-3 fatty acids not found in terrestrial proteins. Therefore, all these biochemical benefits have scientific support for why marine-derived proteins are being investigated for optimal athletic performance. Although diversity in the marine ecosystem is high, only a few organisms have thus far been studied for their bioactive proteins and peptides [8]. These organisms include fish, crustaceans (like crabs, crayfish, lobsters, prawns, and shrimp), invertebrates (like mollusks, sponges, echinoderms, and cnidarians), and various marine microorganisms. Amino acids essential for human metabolic functions can be derived from fish, one of the best natural sources of quality protein. Fish proteins also contain both structural and functional proteins, like collagen, myoglobin, and actin, which are ubiquitously present in tissues like skin, bones, scales, and visceral organs. The proteins and peptides derived from marine resources can be isolated from fish fillets, processing wastages, and coproducts [9]. Besides fish proteins, collagens have also been extracted from other marine animals such as sea cucumbers, mollusks, sponges, crabs, seaweed, and jellyfish [10, 11]. Shellfish such as shrimp, crab, crayfish, and lobster are also excellent sources of protein and functional bioactive peptides. These include antibacterial peptides in blue mussels, shrimp, oysters, and scallops; opioid peptides mostly in mussels and clams; Angiotensin-Converting Enzyme (ACE)-inhibitory in shrimp and crabs; and immunomodulatory in fish, mussels, and scallops. Opioid peptides in shellfish include methionine enkephalins in oysters and clams, dynorphin in the blue mussel (Mytilus edulis), and β-endorphins in shrimp. These inhibit the activity of ACE, which, upon ingestion of the shellfish, works to decrease blood pressure in the cardiovascular system [12-15]. Representative examples include leucine-aspartic acid from clam protein hydrolysates (Meretrix meretrix), isoleucyl-prolyl-proline from oyster (Crassostrea gigas), and isoleucyl-leucine-proline from crab protein hydrolysates (Portunus trituberculatus), whereas valyl-tyrosine (VY) comes from shrimp (Litopenaeus vannamei). In addition, immune-modulatory proteins and peptides in shellfish, such as crustins in crustaceans and hemocyanins in mollusks, increase phagocytosis and induce cytokine production.
Gelatin hydrolysates from sea cucumber, skate skin, jellyfish, and squid skin were shown to lower blood pressure in spontaneously hypertensive rats, which is a risk factor for cardiovascular disease. Fish gelatin hydrolysates/peptides further display an array of bioactivities, which include antioxidant, anemic, immune-modulatory, calcium-binding, mineral chelation, ACE, and blocking the Dipeptidyl-peptidase IV (DPP-IV) inhibition, antibacterial, and anti-hypertensive functions. Such bioactivities were reported for the peptides/hydrolysates from several kinds of fish, including pollack, snapper, Atlantic salmon, cod, herring, hoki, Pacific whiting, and sole in several scientific papers [16, 17]. Antimicrobial Peptides (AMPs) have been discovered in a range of marine animals such as octopus (Octopus vulgaris), sponge (Trichoderma sp.), yellow catfish (Pelteobagrus fulvidraco), mud crab (Scylla paramamosain), marine snail (Cenchritis muricatus), Atlantic cod (Gadus morhua), and oysters (Crassostrea gigas). These AMPs are inexpensive, safe, naturally derived, and highly bioactive; thus, they are promising candidates for various biotechnological applications [18, 19]. Various mollusk species, including bivalves, also possess AMPs. Cysteine peptides are detected in mussels, while defensins and proline peptides are detected in oysters and gastropods. Moreover, other proteins with antimicrobial activity, such as hemocyanins and egg case proteins (Sep-ECPs), can be extracted from gastropods [20]. Recent studies have been increasingly devoted to the isolation of bioactive peptides from diverse marine organisms, such as sponges, tunicates, ascidians, mollusks, fish, and fish products, as well as other marine organisms with prospective anticancer activity [3, 21-23]. Marine organisms such as algae, starfish, mussels, and echinoid worms are also known to be rich sources of anticoagulative peptides [24]. Antioxidative peptides like glutathione and carnosine, which are found in the muscles of mammals, are abundant in marine microalgae [25]. Phycobiliproteins and lectins are the major functional proteins obtained from algae. Phycobiliproteins like phycocyanin and phycoerythrin have been found to have antiviral, anti-inflammatory, and antioxidative effects and can be used as natural food and cosmetics dyes as well as biotechnological markers. Macroalgae lectins like Ulva sp. have been found to be active against bacteria, viruses, tumors, and inflammation through specific glycan binding [26]. All these bioactive molecules have immense potential for the development of various nutraceutical and pharmaceutical formulations.
1.2. Marine Bacterial Dextranases and their Potential Relevance to Performance
Marine bacterial dextranases (EC 3.2.1.11) are hydrolytic enzymes that split the alpha-(1→6) glycosidic bond of dextran, a high-molecular-weight polysaccharide, into low-molecular-weight compounds known as isomaltose-oligosaccharides (IMOs). Marine bacterial dextranases are not only valued for their traditional applications in sugar therapy, oral hygiene, and medicine but are also gaining credence for their applications in sports nutrition. These enzymes, isolated from bacterial cultures adapted to saline and constantly changing marine environments, are remarkably stable in varied pH, salinity, and temperature, thus possessing great promise for the modification of carbohydrates [27, 28]. In sports performance, carbohydrate availability is key to endurance, while the physicochemical characteristics of carbohydrate supplements influence digestion rate, gastric emptying rate, and optimal absorption rate [29]. Using dextranases from marine sources can potentially allow the creation of dextran-based carbohydrates with designed molecular weight profiles to optimize the glucose release rate for prolonged energy. This is similar to the advantages of highly branched cyclic dextrin supplements tested to prolong the time to exhaustion in competitive swimmers [30, 31]. Moreover, dextranase-derived IMOs have proven prebiotic activities as described above to specifically stimulate the growth of lynchpin beneficial microorganisms such as Bifidobacterium and Lactobacillus [32]. There is a growing body of research supporting the importance of the gut microbiome in influencing the athlete's metabolic and inflammatory state and recovery pathways, and this is now further supported by the strategic use of IMOs, made from marine dextranase proteins, in sports nutrition products [33]. In addition, the ability of these enzymes to break down dextran and make a solution less viscous, thereby improving the emptying of carbohydrates in the gut and improving their use in prolonged exercise, when gut issues can be a limiting factor, is a clear benefit in itself and can now be utilized in a new application for IMOs made from marine dextranases. The use of these low molecular weight dextrans, for example, in plasma volume expansion and in the use of drugs, opens up a new range of possibilities for their use in athletes who need rehydration or nutrient delivery in high quantities and in a short interval of time. This potential could open up new pathways for research [29, 34]. Future studies in the clinical area could explore the effect of dextranase-treated carbohydrate supplements on glycemia control and physical performance, while microbiome research could explore the effect of IMO supplements on recovery rate, and finally, the area of biomedicine could explore the safety and regulation of such approaches. In conclusion, the specific combination of catalytic diversity with stability ranks marine bacterial dextranases among the most promising biotechnological approaches available today at the boundary of marine microbiology, carbohydrate biotechnology, and sports biotechnology.
1.3. Marine-Derived Molecules and their Nutritional Values
In recent years, functional and bioactive components isolated from marine organisms, such as bacteria, mollusks, algae, and sponges, have been discovered to possess beneficial health effects and promising therapeutic potential [35]. Marine organisms are known to be rich in novel bioactive components such as peptides, polysaccharides, and fatty acids [35]. Peptides from marine organisms have important functions in basic biological events, such as reproduction, growth, and protection [8]. Marine bioactive peptides can be isolated using solvent extraction or microbial fermentation of protein-based bioactive peptides, generating peptide fragments with three to twenty residues of amino acids [35]. Some studies found purified bioactive peptides from marine organisms to be strong inhibitors of ACE [35]. Seaweed-derived peptides have also been noted to have the potential for inhibiting diabetes and cardiovascular diseases [36], while fish bioactive peptides have been indicated to regulate pathways in blood pressure, lipid, and glucose metabolism and body composition regulation [8]. Some bioactive marine peptides, such as those from Mytilus coruscus, have also been reported as promising candidates compared to traditional antibiotics [19]. More recently, athletes have also commenced supplementing with bioactive peptides derived from marine microorganisms [37]. Bioactive marine peptides have been shown to enhance muscle strength [38], modulate glucose uptake of the muscle tissues [39], and relax the muscles or reduce the soreness resulting from vigorous muscle contraction [40]. Some studies have indicated improvements in muscle strength; however, several of these studies were completed in untrained or recreationally active populations rather than elite athletic populations [41]. These bioactive marine peptides also induce the expression of myosin, actin-binding protein, and tropomyosin-muscle proteins that take part in the adaptation of muscles towards resistance training [42]. Bioactive peptides have also been shown to have the ability to accelerate the translocation of glucose transporters GLUT-4 and GLUT-1 from cytoplasm to the plasma membrane, thus increasing muscle glycogen deposition and anti-stress properties [43, 44]. ACE inhibitory properties of bioactive peptides contribute to their ability to improve endothelial function, expected to benefit endurance exercise performance [45, 46]. Also, the reduction of plasma biomarkers of inflammation and muscle damage induced by bioactive peptides suggests that these compounds might have a role in accelerating musculoskeletal adaptation and recovery by facilitating extracellular matrix remodeling [47, 48]. Branched-chain amino acids, leucine, valine, and isoleucine, are other peptides that have been proven to produce numerous benefits in muscle tissue. These include their capacity to stimulate muscle protein synthesis, improve physical performance and muscle strength, as well as decrease muscle injury induced by exercise [49-51]. Research has confirmed that Branched-Chain Amino Acids (BCAA) supplements are beneficial in enhancing athletic performance. Cheng et al. [52], for instance, demonstrated improved athletic performance in endurance activities in college runners after BCAA supplement administration, while Chen et al. [53] showed decreased central fatigue in taekwondo athletes. Additionally, leucine supplementation has proved to have great therapeutic value in being able to counteract stress-related disorders like burns, traumas, and sepsis, as well as in preventing muscle breakdown. Leucine is found in high concentrations in several species like S. waitei, R. kanagurta, L. rohita, C. mrigala, C. batrachus, and H. fossilis, while isoleucine is prominent in O. mykiss and L. rohita [54]. BCAAs, creatine, and ACE-inhibitory peptides are three bioactive compounds with distinct but complementary roles in physical exercise; the strength of evidence supporting their use differs between mechanism-based studies and human research. Mechanism-based studies have provided evidence on the inhibitory effects of BCAAs against inflammation and fatigue via the mTOR pathway. RCTs have shown the benefits of BCAAs mainly as an anti-exercise phenomenon. Meta-analysis has shown consistent evidence of the inhibitory effect of creatine supplements against serum creatine kinase and the anti-sore effect; however, the findings are not consistent in supporting increased performance capacity or strength gain [55]. On the other hand, the strongest human evidence exists for the supplementation of creatine. This has been established in systematic reviews and large randomized controlled trials to enhance the availability of phosphocreatine and promote ATP replenishment and exercise-related strength and sprint performance (3–5g/day) in both young and older subjects [56, 57]. New evidence has recently emerged for the potential to stimulate the growth of lean mass and to enhance fatigue resistance [56, 57]. While the view has been established that ACE-inhibitory peptides of marine origins are highly mechanistically justified based on animal studies, they are clearly demonstrated to reduce ACE activity and concentrations of angiotensin II, promote signaling mediated by nitric oxide levels, and thus improve physical exercise capacity by improving perfusion and suppressing the accumulation of lactate during endurance exercise (as assessed in animal studies) [58, 59]. Yet, few studies have been undertaken in humans to date, with the majority of trials focusing on reducing blood pressure in hypertensive patients and only a limited number of well-designed controlled trials providing objective assessments of physical performance capacity in athletes, as evaluated by parameters such as VO2 max and time-trial performance [8, 60]. However, ACE-inhibitory marine peptides have demonstrated endothelial and antihypertensive properties in clinical trials; their direct use in trained athletes is currently restricted. The majority of the human trials currently available have been carried out on hypertensive individuals or the general population, and their extension to improve athletic performance in athletes should be interpreted cautiously (Table 1).
| Supplement | Proposed Mechanisms | Animal/ In Vitro Evidence | Human Trial Evidence | Performance Outcomes | Marine Sources |
|---|---|---|---|---|---|
| BCAAs | • mTOR activation• Reduced central fatigue (Trp–BCAA competition)• Anti-inflammatory effects | Strong: reduced inflammation, ↓ lactate, prolonged endurance in rodents [61] | Mixed: ↓ CK and DOMS, faster recovery; inconsistent effects on strength/time-trials [62] | Primarily recovery-focused; limited direct ergogenic benefit [61] | Fish protein hydrolysates (e.g., tuna, salmon), shrimp, squid, seaweed-derived peptides |
| Creatine | ↑Phosphocreatine storage• Faster ATP resynthesis• Improved neuromuscular function | Strong: consistent increases in PCr, ATP buffering, muscle hypertrophy in rodents and cell studies [56] | Robust: ↑ strength, ↑ power, ↑ repeated sprint capacity; lean mass gains well-documented [57] | Best-supported ergogenic aid: strength, power, sprint performance [56, 57] | Fish (e.g., herring, salmon), shellfish (e.g., crab, shrimp) |
| ACE-Inhibitory Peptides (marine-derived) | • Inhibit ACE → ↓ Ang II• ↑ Endothelial NO signaling• Improved perfusion and anti-fatigue | Strong: antihypertensive, vasodilatory, anti-fatigue (swimming tests, ↓ lactate in mice) [58, 63] | Limited: mostly BP/endothelial outcomes in hypertensive adults; no clear athlete RCTs yet [8, 60] | Promising but unproven for athletes; potential for endurance gains via vascular function [63] | Fish (e.g., sardine, tuna, salmon), shrimp, mussels, sea cucumber |
Other marine species, from the tunas, mackerels, emperor fish, silky sharks, to crustaceans like lobsters and crabs, have also been reported to have substantial composition of the leucine, isoleucine, and valine amino acids. Recently, it was estimated that a serving of spiny lobster or spanner crab can provide around 60–67% of the day's needs for valine, leucine, and isoleucine, while a serving of fish was claimed to be sufficiently adequate for more than 100% of the daily recommended intake for essential amino acids [64]. Other ergogenic values contributed by marine foods may be attributed to high beta-alanine, creatine, and hydroxymethylbutyrate (HMB) from fish and algae sources [65, 66]. Beta-alanine supplementation has been reported to increase exercise duration until fatigue, increase time to exhaustion, and improve power output during resistance exercises [67, 68]. Creatine supplementation decreases fatigue and improves energy stores during intense exercise [69]. Athletes using HMB supplements see numerous performance benefits, including increased anaerobic peak and average power, decreased lactate accumulation following anaerobic exercise, as well as body composition changes resulting from fat loss and lean muscle gain. HMB also appears to reduce overreaching by dampening stress hormone responses [70, 71]. Marine-derived antioxidants may also boost immune function and athletic performance by protecting muscle tissue from oxidative damage [72]. Antioxidant supplementation has been reported to lower oxidative stress in young soccer players, as demonstrated by lower glutathione-to-oxidized-glutathione ratios and improved markers of lipid peroxidation, such as malondialdehyde [73]. In addition, antioxidant supplementation aids athletes' training at altitude by lowering red blood cell deformity [74]. When taken before and/or during physical activity, antioxidants have also demonstrated the ability to decrease recovery time as well as postpone fatigue [75]. Marine-derived peptides that have ACE inhibitory activity may also have a positive effect on cardiovascular activity during physical exercise. During physical activity, heart rate and blood pressure increase to ensure proper oxygenation of active muscles through autoregulation. Marine-derived ACE inhibitory peptides may have a regulatory effect on this aspect by inhibiting angiotensinogen transformation to angiotensin II [76]. This will increase vasodilation and ensure optimal blood pressure regulation. This mechanism allows for reduced cardiac overexertion during intense physical activity. Apart from enhanced cardiac activity, marine-derived ACE inhibitory peptides may have positive effects on kidney function by lowering glomerular pressure and mitigating kidney overexertion during prolonged physical activity. A decrease in blood pressure after physical activity occurs when vasodilatory hormones such as nitric oxide are secreted to ensure enhanced blood circulation [77].
Thus, the molecules for marine peptides have a great potential as a natural sports supplement. However, despite the promising bioactivities of these molecules, the use of marine peptides as a sports supplement has yet to be fully explored. Research indicates that administering conventional pharmaceutical ACE inhibitors to athletes yields low levels of synergistic effects, warranting further investigation into the efficacy of the natural marine peptide compound [78].
1.4. The Potential Roles of Dextranases, Collagenases, and Fucoidanases
Marine environments, characterized by high salinity, pressure, and temperature variations, supported the evolution of microorganisms and invertebrates that produce stable, catalytically diverse enzymes. Dextranases, collagenases, and fucoidanases from marine organisms are valuable not only for their biotechnological and medical applications but also for their emerging use as assistive aids to enhance human athletic performance. Their special biocatalytic properties can be considered as the development of a new toolbox in the study of sports sciences in energizing and recommending recovery and overall robust health. Dextranases (EC 3.2.1.11) are specifically known to split the α-(1→6) glycosidic bonds of dextran into IMOs. Marine dextranases have better properties than those from terrestrial organisms for industrial use, as they tolerate higher salt concentrations, pH, and temperature [27, 79]. Such properties make them optimal for sports nutrition applications, such as gels and sports drinks. As carbohydrates are the primary component of sports nutrition, glycogen depletion, particularly during prolonged activities, can cause fatigue, making it essential to supplement it with externally sourced compounds [29]. The structure of ingested carbohydrates affects the different absorption rates and their glycemic index. The strategy of using dexterase-treated carbohydrates to release glucose and drive hydrolysis to oligosaccharides in sports applications, hence avoiding “sugar crashes,” may be conceptualized with highly branched cyclic dextrin when this increases the time to exhaustion for swimmers compared with glucose supplementation [30]. These derived compounds also exert a prebiotic effect, promoting the growth of healthy gut microorganisms such as Bifidobacterium and Lactobacillus [32]. This becomes even more important in view of the known relationship between the gut-muscle axis and performance, modulating nutrient absorption, systemic inflammation, and stress response [33]. The marine dextranases, therefore, might provide a dual benefit in terms of an improvement in carbohydrate metabolism and gut health. Collagenases (EC 3.4.24.3) catalyze the cleavage of collagen into smaller peptides. The marine collagenases from bacteria and fish yield Bioactive Collagen Peptides (BCPs), and these peptides show antioxidant, anti-inflammatory, and tissue repair effects [80, 81]. Peptides can be directly utilized in repairing the musculoskeletal system due to micro-trauma caused in muscle, tendons, and joints with intense exercise. The collagen peptides trigger extracellular matrix remodeling and connective tissue synthesis, and there is clinical evidence suggesting a reduction in joint pain and improvement in mobility in actively working people with the use of collagen hydrolysates [82]. These antioxidant peptides could also neutralize exercise-induced free radicals, reducing oxidative stress, a major factor causing fatigue and delayed recovery, thus mitigating muscle soreness and inflammation. In this light, marine collagenases represent an interesting opportunity not only in the context of recovery after injury but also as nutritional supplements contributing to the maintenance of joint integrity and accelerated post-workout regeneration. Fucoidanases (EC 3.2.1.44) hydrolyze the fucoidan sulfated polysaccharide from brown seaweed into bioactive oligosaccharides possessing anticoagulant, antioxidant, immunomodulatory, and anti-fatigue properties [63, 83]. In relation to athletes, these features are particularly relevant, since enhanced circulatory efficiency is crucial for endurance, and fucoidan-derived oligosaccharides with anticoagulant potential may improve blood circulation and oxygen delivery. Anti-fatigue effects in animal studies have been supported by inhibitions in blood lactate concentration and enhanced swimming ability [84]. In addition to that, immunomodulation is an important point to note, especially with respect to an immunosuppressed condition brought on by rigorous training schedules, placing athletes at greater risk for diseases due to increased susceptibility to infections. The elongated swimming time and the lower level of blood lactate have been the primary indicators of anti-fatigue effects in animal experiments. However, there are not many controlled human trials in sporty groups at the moment [85]. Fucoidan oligosaccharides have already shown potential for activating immune cells and enhancing resilience to stress. In other words, these three enzymes create a basis or platform to establish marine enzymology because they are utilized in sports. These enzymes, such as dextranases, are specifically designed to function in fueling systems and gut processes. Then there are those, like collagenases, focusing on activating processes to repair any damaged tissues, using antioxidant properties. Fucoidanases activate processes to enhance circulation, fatigue resistance, and immune functions.
1.5. Collagen Peptides in Athletic Performance: From Molecular Nutrition to Clinical Outcomes
Collagen Peptides (CPs) have a proven role in nutritional therapy, and evidence has appeared, coming from trials, mechanisms, and systemic analyses, for their effectiveness in strength augmentation, healing, and injury prevention. Collagen is recognized to be the principal structural protein found within tendons, ligaments, and cartilage. Heavy exercise impacts this tissue adversely by causing extensive micro-damage, thus impeding performance. CP is known to contain active peptides rich in proline, hydroxy-proline, and particularly, glycine. They are produced from Collagen, a biomaterial of considerable commercial efficacy. The commercial name for Collagen peptide is C-collagen peptide, which is an acronym for Collagen peptide. These agents act as precursors in collagen production and promote fibroblast function, thus helping in improving tissue remodeling when subjected to mechanical stress. Clinical evidence supports CPs improving musculoskeletal responses directly. For example, in a 12-week double-blind randomized controlled trial, it was observed that runners who received 15 g/day of specific collagen peptides demonstrated better one-hour time trial and speed of lactate threshold values as opposed to the placebo group, thus suggesting improvement in endurance training efficiency [86]. Moreover, resistance training experiments in young participants have uniformly demonstrated greater patellar tendon stiffness, cross-sectional area, and explosive power in response to CP administration as opposed to iso-caloric placebo conditions [87, 88]. These particular tendon modifications play a critical role in their own right, as an increase in tendon stiffness enhances force transmission and power output and also prevents strain injuries. Mechanistic work supports these results. Shaw et al. (2017) affirmed in a study with human subjects that ingestion of vitamin C-enriched gelatin one hour prior to high-impact loading elevated blood markers of collagen synthesis two-fold, and timing the ingestion of CP with exercise has a key role in enhancing extracellular matrix remodeling [89]. In addition to these advantages, CP facilitates recovery following exercise through attenuation of muscle damage and soreness. A clinical trial with young, healthy males suggested that CP supplementation attenuated muscle soreness and fatigue and accelerated recovery of muscle strength following standardized muscle damage induced by exercise [90]. A recent trial in athletes undergoing concurrent training in 2024 also affirmed in an RCT that particular collagen peptides attenuated serum markers of muscle stress [48]. These effects are probably due to an improvement in connective tissue healing and the intrinsic antioxidant activity of the collagen-derived peptides, which may neutralize free radicals induced by exercise. CPs also exert an important preventive and rehabilitative role in sports medicine. An early 24-week RCT in 147 college athletes demonstrated that collagen hydrolysate significantly reduced activity-related joint pain, with the most robust effect among those engaged in high-impact activities [82]. More recent trials in patients with chronic Achilles' tendinopathy confirm that daily supplementation with particular collagen peptides, in association with a strengthening protocol, significantly improved pain and functional outcomes versus placebo, with an ex novo confirmation of their therapeutic value [91]. Systematic reviews are continually adding to the basis of this evidence. For instance, a conclusion drawn by Khatri et al. in 2021 states that CPs are effective in decreasing joint pain and increasing joint function both in athletes and non-athletes alike [48]. A more contemporary review, undertaken by Bischof et al. in 2024, included trials up to 2023 and suggested a marked and homogenous effect for CP on tendon stiffness, recovery indices, and self-reported pain, albeit with a degree of variability in studies for factors such as protocol and dosing, and source of ingested collagen protein [92]. From a clinical perspective, the best regimen for athletes is becoming clear and involves 10-15 grams of hydrolyzed protein administered 30-60 minutes before tendon-bearing exercise and concomitantly with vitamin C, for a course of no less than 8-12 weeks. The benefits to specific sports are clear: in endurance sports, CPs may enhance running economy and lower the chance of tendon damage; for strength and power sports, they improve tendon stiffness and force transmission; for high-impact athletes, they reduce joint pain and overuse injuries and may quicken the return-to-play timeline. In conclusion, clinical, mechanistic, and translational data strongly place collagen peptides as a specific, evidence-based nutritional intervention. They act as a very active bridge between nutrition at the molecular level and athletic performance by enhancing connective tissue adaptation, reducing injury risk, and hastening recovery in endurance, strength, and hybrid disciplines.
1.6. Comparative Advantages of Bioactive Peptides from Marine, Animal, and Plant Sources
Bioactive peptides may be sourced from a variety of animal and plant species. Every potential source of bioactive peptides has its own merits and demerits. Biopeptides sourced from animal species such as meat, eggs, and milk are currently in use as performance-enhancing supplements to exploit their essential amino acid profile and established role in the muscle recovery process [93, 94]. Use of these types of biopeptides may be associated with some shortcomings, such as potential allergies, increased intake of saturated fats, concerns regarding the sustainability of food resources, and the possibility of reduced peptide bioactivity after processing or heating [95]. Recently, the use of peptides from plants has become more popular in the scientific community due to sustainable factors in particular, and increasingly as a consequence of the popularity of a vegetarian diet [96]. Although plant peptides have several advantages over animal proteins, such as being more ecologically sound and humane, their use in sports nutrition might be limited by lower digestibility and a lack of a complete essential amino acid profile compared to animal proteins [97, 98]. Furthermore, the presence of certain antinutritional factors in plants, phytates and tannins, that play a detrimental role in the absorption of amino acids, might affect the efficacy of the use of peptides from plants [99]. Seafood peptides have many good characteristics that would make them specifically desirable in a sport-active group of people. They are all very digestible and have a quick absorption rate and a complete amino acid structure suitable for muscle function support [93]. Beyond nutritional value, many marine peptides possess additional bioactivities, including antioxidant, anti-inflammatory, and anti-fatigue effects, each of which directly addresses the physiological stresses associated with intense or prolonged exercise [100-102]. These are often related to a relatively low molecular weight and distinctive amino acid sequences, reflecting adaptation to the challenging conditions in the marine environment [103, 104]. Such benefits notwithstanding, application of marine-derived peptides is not without its challenges. Variability in raw material composition, possible sensory drawbacks such as taste or odor, as well as higher extraction and purification costs, can stand in the way of large-scale application [3, 105]. However, recent progress within enzymatic hydrolysis techniques, purification technologies, and the use of marine processing by-products serves to overcome these limitations while enhancing both viability and sustainability of marine peptides in sports nutrition applications [106, 107]. Pharmacological and Biochemical Outcomes of Marine-Derived Compounds have been summarized in Table 2.
| Outcome Category | Key Biochemical / Pharmacological Effects | Underlying Mechanisms | Exercise / Performance Relevance | Representative Marine Sources |
|---|---|---|---|---|
| Muscle Protein Metabolism | ↑ Muscle protein synthesis; ↓ proteolysis | Activation of mTOR signaling; increased availability of Leu-rich peptides | Supports muscle hypertrophy, strength gains, and training adaptation | Fish protein hydrolysates (salmon, tuna), shrimp, squid [93, 94, 108] |
| Anti-Fatigue Effects | ↓ Blood lactate; ↑ glycogen preservation | Improved mitochondrial efficiency; reduced central fatigue | Delays fatigue during prolonged or high-intensity exercise | Fish peptides, sea cucumber, algae-derived peptides [109, 110] |
| Antioxidant Defense | ↓ ROS; ↑ SOD, CAT, GPx activity | Free radical scavenging; upregulation of endogenous antioxidant enzymes | Limits exercise-induced oxidative stress and muscle damage | Fish skin/collagen peptides, shellfish, microalgae [104, 111, 112] |
| Anti-Inflammatory Activity | ↓ IL-6, TNF-α, CRP; ↓ CK | Modulation of NF-κB and COX-2 signaling | Enhances recovery and reduces DOMS | Fish hydrolysates, mussels, shrimp [20, 100, 113] |
| Vascular Function | ↑ NO bioavailability; ↓ ACE activity | ACE inhibition; improved endothelial function | Improves oxygen and nutrient delivery during endurance exercise | Sardine, tuna, salmon peptides [114-116] |
| Energy Metabolism | ↑ Fat oxidation; improved glucose uptake | Activation of AMPK; enhanced metabolic flexibility | Supports endurance capacity and metabolic efficiency | Fish and algae-derived peptides [117-119] |
| Connective Tissue Support | ↑ Collagen synthesis; improved tendon integrity | Supply of Gly-Pro-Hyp peptides; fibroblast stimulation | Supports joint and tendon health under training load | Fish collagen, fish skin, scales [48, 120, 121] |
| Gut Absorption and Bioavailability | Rapid absorption; high peptide stability | Low molecular weight peptides; resistance to GI degradation | Faster nutrient delivery post-exercise | Marine protein hydrolysates [106, 107] |
1.7. Future Implications and Direction in Nutraceutical Use in Athletes
Globally, the concern about the performance of elite sports is observed in the pursuit of developing innovative nutrition therapies. Optimum power is needed in moderate to highly intense sports, and the process requires a far more significant measure of lean muscle mass and reduced body fat. Nutraceuticals can be quite effective to the extent that they can stimulate the body towards avoiding effective nutrition challenges while demanding better performance as well [122]. However, intense training can stimulate negative physiological responses such as the disruption of the immune system and the occurrence of inflammation and oxidative stress. Therefore, coaches and players must make optimum assessments specific to each individual's physiological demand. This can be achieved by making use of effective scientific assessments based on key markers, with a deeper look, for example, into the biological fluids such as the impacts of the elements found in the blood, urine, and feces. The obtained data could then be used to direct proper functional supplement usage and sport nutrition practices [123, 124]. Personalized sports nutrition plans could also be created in different ways, including combining various nutrients in one delivery form or using different delivery forms that contain varied nutrient profiles. Personalized formulas are meant to address the personal physiological and performance requirements of different individuals (Fig. 2).
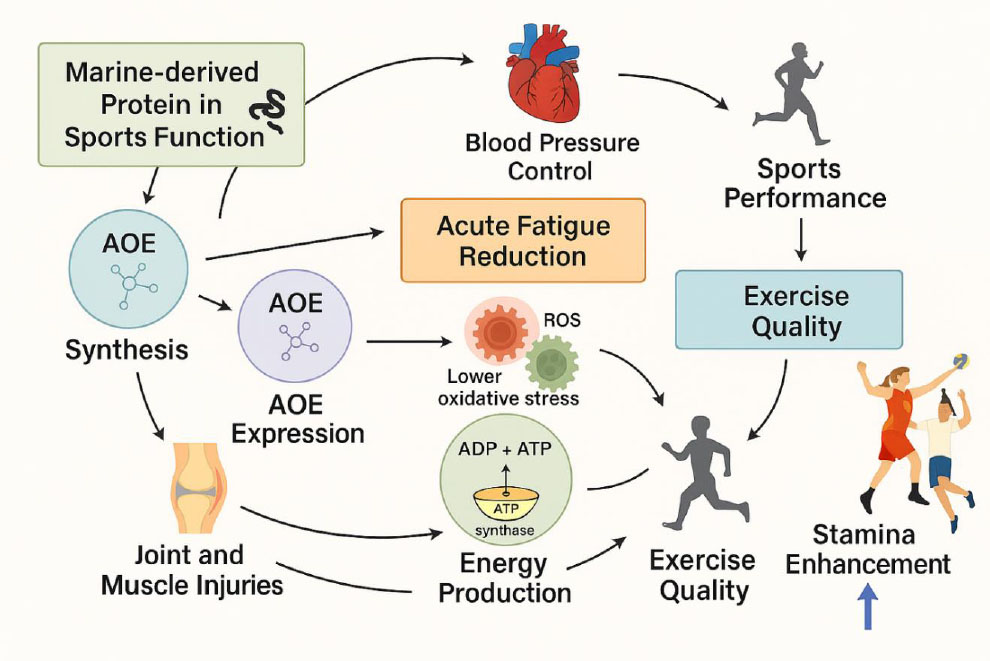
Mechanisms of action of marine-derived proteins and peptides on athletic performance.
The ergogenic effect of marine-derived compounds is accomplished by several mechanisms working in tandem: (i) Vascular Regulation, whereby active peptides with ant Angiotensin-Converting Enzyme (ACE) activity cause vasodilation via modulation of the renin-angiotensin complex; (ii) Antioxidant Defense, whereby peptides counteract oxidative stress generated by exercise by scavenging ROS and fortifying antioxidant defenses; (iii) Metabolism of Energy, in which dextranases, enzymes of marine origin, adapt carbohydrate breakdown to supply continuous energy; and (iv) Musculoskeletal Protection, whereby collagen peptides provoke the manufacture and repair of tendons and ligaments. Such mechanisms illustrate the promising role of marine-derived compounds as new tools in increasing endurance, strength, and performance recovery [79, 82, 91]. The ability of compounds from the sea to help in overcoming health problems caused by obesity, such as inflammation, oxidative stress, diabetes, and dyslipidemia, has attracted a great deal of attention in recent years. A variety of bioactive substances from the sea are presently under investigation for their medicinal values, which have proved promising in overcoming different metabolic disorders. Marine-based food supplements are a rich source of highly bioactive compounds like proteins, peptides, carotenoids, glucosamine, minerals, and omega-3 fatty acids. Marine-based food supplements make a valuable nutritional source for athletes due to their ability to increase endurance and counteract the different nutritional requirements of a person who is involved in intense exercise and activity [122]. A particular benefit of omega-3 fatty acids is their proven efficacy in muscle function and metabolism in relation to physical activity. These properties of omega-3 fatty acids include antioxidant and anti-inflammatory effects, as well as an ability to induce rapid cell regeneration, thus improving muscle recovery and function. Most studies are conducted on clinical or non-athletic groups, despite the fact that some human studies have revealed a reduction in the indicators of oxidative stress. At present, there is not enough information available to determine a direct correlation between these effects and the results of enhanced athletic performance [122-126]. On the other hand, the improvement in muscle functions, as well as the reduction in muscle injury, has been shown to be related to the consumption of marine bioactive peptides [127]. Given this context, it can thus be noted that marine products have the potential to serve as sustainable, eco-friendly alternatives for sports supplements. Given the varied nutritional content and distinct bioactive compounds found in these products, they certainly hold more potential than other sports supplements. Moving on, a more precise explanation of marine peptides would require a comparison with other, more traditional, land-based ingredients, such as gelatin derived from cows or chickens. While these terrestrial gelatins are in widespread use, they are generally composed of much larger peptide fragments that are poorly soluble and less available than their marine-derived counterparts [81]. With their much lower molecular weight profiles, the marine collagen peptides are incomparably more absorbable and hence are considered particularly promising for athletic use. The physiological outcomes importantly depend on dosage, as clinical studies demonstrated that 15 g/day for 12 weeks improved running endurance [87], whereas a treatment with 5 g/day, together with strengthening exercises, allowed tendon repair in tendinopathy patients [91]. Moreover, the specific cardiovascular effect of marine peptides should be described. The mechanism of action of the ACE inhibitory variants involves the inhibition of angiotensin II and the potentiation of nitric oxide. This mechanism is known to increase cardiac efficiency as a physiological phenomenon rather than as a pathological burden. Taken together, these observations point out the functional advantage of marine peptides and the need to apply them practically based on the consideration of the respective dosage and effects.
1.8. From Evidence-Based Ergogenic Application to Biological Believability
Although marine peptides do have the potential to activate mTOR, inhibit ACE, activate antioxidant pathways, and modulate extracellular matrix, these biological processes should not be considered direct markers of athletic performance. Most of the data that are currently available have been generated in vitro, in animal models, or in human subjects with cardiovascular or metabolic disease. There have been few well-controlled randomized clinical trials evaluating direct performance outcomes such as VO2max, time to exhaustion, sprint speed, or maximal strength in athletic populations, despite the biological plausibility of these findings. Although ACE inhibitory peptides, fucoidan analogs, and dextranase-derived carbohydrates are biologically plausible but not adequately proven in athletic populations, collagen peptides and creatine are the marine compounds with the most evidence for human performance. As depicted in Table 3, the rapidly growing worldwide market for marine-source proteins in the sports nutrition and functional food sectors is another indication of the growing interest in these proteins.
| Marine-Derived Protein Segment | Market Value (Latest Estimate) | Forecast Growth | Key Applications |
|---|---|---|---|
| Marine-derived proteins (overall market) | ~USD 8.9 billion | Expected to reach ~USD 17.5 billion by 2032 (CAGR ≈ 8.8%) | Functional foods, sports nutrition, nutraceuticals [58] |
| Marine protein ingredients | ~USD 4.8 billion | Projected ~USD 7.8 billion by 2035 (CAGR ≈ 4.9%) | Protein supplements, food fortification [92] |
| Marine collagen | ~USD 1.26 billion | Projected ~USD 2.82 billion by 2034 | Joint health, muscle recovery, skin health [92] |
| Marine collagen peptides | ~USD 1.76 billion | Expected to exceed ~USD 5 billion by 2035 | Sports supplements, functional foods [111] |
CONCLUSION
Despite the large potential of sea-derived proteins and peptides as an ergogenic agent for athletes, there exist a number of important drawbacks. Firstly, the problems of oral bioavailability and stability during body metabolism remain major challenges, as enzymes break down many biological peptides before they reach their target sites. Secondly, much of the mechanistic evidence has come from in vitro or animal studies, whereas human trials are often performed on a clinical or non-athletic population. Well-powered randomized controlled trials are required to verify direct ergogenic effects in a trained athletic population. Finally, many factors pose major constraints, since there may be significant differences in extraction and purification procedures, and many protocols are needed for market approval. Finally, scalability and sustainability are emerging as important areas, given that available marine resources are limited and large-scale production must occur in a responsible manner that prevents harming the ecosystems. Thus, although marine compounds such as collagen hydrolysates, ACE-inhibitory peptides, and oligosaccharides derived from the enzyme dextranase have very promising applications in athletics and endurance, their development in sports nutrition requires careful validation and regulation.
AUTHORS’ CONTRIBUTIONS
The authors confirm contribution to the paper as follows: Study conception and design: Data collection: N.D.A.H. and F.M.: Analysis and interpretation of results: All authors reviewed the results and approved the final version of the manuscript.
LIST OF ABBREVIATIONS
| ACE | = Angiotensin-converting enzyme |
| AMPs | = Antimicrobial peptides |
| IMOs | = Isomaltose-oligosaccharides |
| BCAA | = Branched-Chain Amino Acids |
| HMB | = Hydroxymethylbutyrate |
| BCPs | = Bioactive collagen peptides |
| CPs | = Collagen peptides |
ACKNOWLEDGEMENTS
Declared none.

